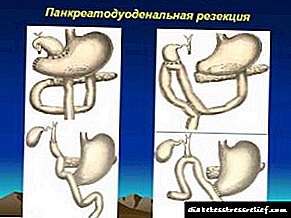
Kolesterolaren biosintesia eta haren biokimika - Diabetesa
Zalantzarik gabe, kolesterola jendarte osoarentzako lipido ezagunena da, oso ezaguna da odol kolesterol handia eta giza gaixotasun kardiobaskularrek duten maiztasunagatik. Kolesterolen zelula-mintzak osagai gisa eta hormona esteroideen eta behazun azidoen aitzindari gisa arreta gutxiago jarri da. Kolesterola animalia askorentzat beharrezkoa da, gizakiak barne, baina ugaztunen elikagaietan duen presentzia aukerakoa da. Gorputzeko zelulek beraiek aitzindari soiletatik sintetiza dezakete.
27 karbono konposatu honen egiturak bide bat konplexua iradokitzen du bere biosintesia egiteko, baina karbono atomo guztiak aitzindari bakar batek ematen ditu: azetatoa. Isopreno blokeak azetatetik kolesterolera bitartekaririk garrantzitsuenak, lipido natural askoren aitzindariak dira, eta isopreno blokeak polimerizatzen dituzten mekanismoak antzekoak dira bide metaboliko guztietan.
Kolesterolaren biosintesia azetatoaren bideko fase nagusiak aztertzen hasten gara. Ondoren, kolesterola odoletik garraiatzeaz, zelulek bere xurgapenaz, kolesterolaren sintesi erregulazio normala eta erregulazioa xurgatu edo garraiatzea eragiten duten kasuetan eztabaidatzen dugu. Orduan, kolesteroletik datozen beste substantziak aztertzen ditugu, hala nola, behazun azidoak eta hormona esteroideak. Azkenik, konposatu asko osatzeko bide biosintetikoen deskribapena - isopreno blokeen eratorriak, zeinetan kolesterol sintesiarekin hasierako fase ohikoak diren, biosintesian kondentsazio isoprenoideen aparteko aldakortasuna ilustratuko da.
Kolesterola azetil-CoA-tik sortzen da lau fasetan
Kolesterola, kate luzeko gantz-azidoak bezala, azetil-CoAz eginda dago, baina muntaketa eredua guztiz bestelakoa da. Lehen esperimentuetan, metil edo karboxil karbono atomoan 14 C-rekin etiketatutako azetatoa gehitu zitzaien animaliei. Etiketa bi animalien taldeetatik isolatuta dagoen kolesterolean banatuta (21-32 irudia), kolesterolaren biosintesi fase entzimatikoak deskribatu ziren.
Fig. 21-32. Kolesterolaren karbono atomoen iturria. Metil karbonoarekin (beltza) edo karboxil karbonoarekin (gorria) etiketatutako azetato erradioaktiboa erabiliz esperimentuak identifikatu ditu. Egitura kondentsatuan, A eta D hizkiek adierazten dituzte eraztunak.

Sintesia lau etapatan gauzatzen da, irudian ikusten den moduan. 21-33: (1) hiru azetato hondakinen kondentsazioa mevalonatoaren sei karbono bitarteko bat osatzeko, (2) mevalonato bihurtzeko isopreno blokeak bihurtzeko; (3) bost karbono isoprenoko sei unitate polimerizatzea 30 karbono eskaleno lineal osatzeko, (4) eskalenoen zikloia osatzeko. esteroide nukleoaren lau eraztunak, eta jarraian hainbat aldaketa (oxidazio, kentzea edo talde metilen migrazioa) kolesterola sortzearekin batera.
Fig. 21-33. Kolesterolaren biosintesi orokorraren argazkia. Testuaren lau fase aztertzen dira testuan. Squalene-ko isopreno blokeak marra marradun gorriekin markatuta daude.

Etapa (1). Azetatoaren mevalonatoaren sintesia. Kolesterolaren biosintesi lehen etapak bitarteko produktu bat eratzea dakar mevalonate (21-34. Irudia). Bi acetil CoA molekulak kondentsatzen dira acetoacetyl CoA emateko, hirugarren acetyl CoA molekula kondentsatzen duten sei karbono konposatu bat osatzeko β-hidroxy-β-metilglutaryl-CoA (HM G -CoA). Bi lehen erreakzio hauek katalizatuta daude thiolase eta NM G -CoA sintasa, hurrenez hurren. cytosolic NM G-CoA sintasa Bide metaboliko hori ezberdina da isoenzima mitokondrialaren arabera, zeinak NM G -CoAren sintesia katalizatzen duen ketona-gorputzak eratzean (ikus 17-18. Irudia).
Fig. 21-34. Acetil-CoA-tik mevalonatoaren eraketa. Azetil-CoAren C-1 eta C-2 mevalonato iturria arrosez nabarmentzen da.

Hirugarren erreakzioak prozesu osoaren abiadura mugatzen du. Bertan, NM G -CoA mevalonatera murrizten da, eta horretarako, NА D PH molekula bakoitzak bi elektroi ematen ditu. HMG-CoA reductase - ER leuna duen mintz integraleko proteina. Geroago ikusiko dugun moduan, kolesterolen eraketa metabolikoaren bide metabolikoa erregulatzeko puntu nagusia da.
Etapa (2). Mevalonato bi isopreno aktibatu bihurtzea. Kolesterolaren sintesiaren hurrengo fasean, hiru fosfato talde transferitzen dira ATP molekuletatik mevalonatura (Fig. 21-35). C-3 mevalonatoan hidroxilo taldearekin lotzen den fosfatoa, tarteko 3-fosfofosfatrofonvalonatoa alde batera utzitako talde ona da, hurrengo urratsean bi fosfato horiek eta aldameneko karboxilen taldea alde batera utzita, lotura bikoitza osatzen dute karbonoaren bost karbonoan ∆ 3 -isopentenil pirofosfatoa. Hau aktibatutako bi isoprenes lehenengoa da - kolesterol sintesian parte hartzaile nagusiak. Δ 3-isopentenilpifosfatoaren isomerizazioak bigarren isopreno aktibatua ematen du dimetilalil pirofosfatoa. Landareetako zelulen zitoplasman isopentenil pirofosfatoaren sintesia hemen deskribatutako bidearen arabera gertatzen da. Hala ere, landare kloroplastek eta bakterio askok mevalonatetik independentea den bidea erabiltzen dute. Bide alternatibo hau ez da animalietan aurkitzen, beraz, erakargarria da antibiotiko berriak sortzerakoan.
Fig. 21-35. Mevalonato bihurtutako isopreno bloke aktiboetan bihurtzea Aktibatutako sei unitateak squalene osatzeko konbinatzen dira (ikus 21-36 irudia). 3-fosfo-5-pirofosfomevalonatoen talde uzten arrosak nabarmentzen dira. Parentesi karratuetan bitarteko hipotetikoa da.

Etapa (3). Sei isopreno aktibatutako unitateen kondentsazioa squalene osatzeko. Isopentenil pirofosfatoa eta dimetilalil pirofosfatoa gaur egun buruko isatsaren kondentsazioa jasaten ari dira. Horietan pirofosfatoen talde bat mugitzen da eta 10 karbono kate bat eratzen da - geranil pirofosfatoa (21-36 irudia). (Pirofosfatoa burura lotzen da.) Geranil pirofosfatoak buruko isatsaren kondentsazioa izaten du isofentenil pirofosfatoarekin, eta 15 karbono bitarteko forma ditu farnesil pirofosfatoa. Azkenean, farnesil pirofosfatoaren bi molekulek "buruarekin" bat egiten dute, bi fosfato-taldeak kendu egiten dira. squalene.
Fig. 21-36. Squalene eraketa. 30 karbono atomo dituen squalene egitura isopreno (bost karbono) blokeek aktibatzen dituzten ondoz ondoko kondentsazioetan gertatzen da.

Bitartekari horien izen arruntak lehen isolatu ziren iturrien izenetatik datoz. Geraniolek, arrosaren olioaren osagaiak, geranio zaporea du, eta akarnas farnesaren koloreetan aurkitutako farnesolak bailarako usain lilia du. Landare natural usain asko isopreno blokeetatik sortutako konposatuei dagozkie. Squalene, marrazo gibetik (Squalus espeziea) isolatuta dago lehenik, 30 karbono atomoz osatuta dago: 24 atomo kate nagusian eta sei atomo metalikoen ordezkoetan.
Etapa (4). Squalene nukleoa esteroide baten lau eraztunetan eraldatzea. Fig. 21-37 garbi ikusten da squalene katearen egitura, eta esterolak ziklikoak direla. Esteroide guztiek esteroide nukleoa osatzen duten lau eraztun kondentsatuak dituzte eta denak C-3 atomoan hidroxil taldea duten alkoholak dira, hortik dator izen ingelesa sterol. Ekintzapean squaloen monooxigenasa O-tik oxigeno atomo bat gehitzen zaio squalene katearen amaieran 2 eta epoxidoa eratzen da. Entzima hau beste funtzio mistoko oxidasa da (21-1 gehigarria), NADPH-ek O oxigenotik atomoko beste oxigeno bat murrizten du 2 H-ra2 O. Produktuaren lotura bikoitzak squalene-2,3-epoxidoaren ordenatuta, erreakzio oso koherentea epoxidoaren eskaleno kateak egitura ziklikoa bihurtu dezake. Animalia-zeluletan ziklizazio horrek eraketa dakar lanosterol, esteroide nukleoaren ezaugarri diren lau eraztun ditu. Ondorioz, lanosterola kolesterola bihurtzen da gutxi gorabehera 20 erreakzio sortaren bidez, metal talde batzuen migrazioa eta beste batzuk kentzea barne hartzen ditu. Ezagutzen artean zailenetakoa den biosintesi bide harrigarri honen deskribapena Conrad Bloch-ek, Theodore Linen-ek, John Cornfort-ek eta George Popiak-ek egin zuten 1950eko hamarkadaren amaieran.
Fig. 21-37. Eraztun itxiak squalene lineala kondentsatutako esteroide nukleo bihurtzen du. Lehenengo etapa funtzio mistoa (monooxigenasa) duen oxidasa batek katalizatzen du, eta horren cosubstratoa N AD PH da. Produktua epoxidoa da, hurrengo fasean zelula esteroide bat osatzeko. Animalia-zeluletan erreakzio horien azken produktua kolesterola da: beste organismo batzuetatik haratagoko antzerkiak desberdinak dira.


Kolesterola animalien zelulen ezaugarri esterol bat da, landareek, onddoek eta protistek oso antzeko esterolak sortzen dituzte.
Sintesi bide bera erabiltzen dute squalene-2,3-epoxidoa lortzeko, baina gero bideak pixka bat desbideratzen dira, eta beste esterolo batzuk eratzen dira, esaterako, sigmasterol landare askotan eta ergosterola onddoetan (Fig. 21-37).
21-1 adibidea Squalene-sintesiarako energia-kostuak
Zein dira squalene molekula bat sintetizatzeko energia kostuak (ATP molekulak bezala adierazten dira)?
Erabakia. Azetil-CoAren eskalenoen sintesian, ATP mevalonatoa aktibatutako isoprene squalene aitzindari bihurtzen denean gertatzen da. Isopreno aktibatutako sei molekula behar dira squalene molekula eraikitzeko, eta hiru ATP molekula behar dira aktibatutako molekula bakoitza sortzeko. Orotara, 18 ATP molekula gastalen molekula baten sintesian gastatzen dira.
Kolesterola gorputzean
Ornodunetan, kolesterol kopuru handia sintetizatzen da gibelean. Bertan sintetizatutako kolesterolaren zati bat hepatozitoen mintzetan txertatzen da, baina batez ere bere hiru forma hauetako batean esportatzen da: kolesterol biliar (biliar), azido biliar edo kolesterol esterrak. Hezur azidoak eta haien gatzak kolesterolaren eratorritako hidrofiloak dira, gibelean sintetizatzen direnak eta lipidoen digestioan laguntzen dutenak (ikus 17-1 irudia). Kolesterolaren esterrak gibelean eratutako ekintzaz azil-CoA-kolesterola-akiltransferasa (ACAT) Entzima honek azido gantz baten hondakina A koentzimatik kolesterolaren talde hidroxilera transferitzea katalizatzen du (Fig. 21-38), kolesterola forma hidrofoboagoa bihurtuz. Lipoproteinaren partikuletan jaritako kolesterola esterrak beste ehunetara eramaten dira kolesterola erabiliz edo gibelean gordetzen dira.
Fig. 21-38. Kolesterolaren sintesia. Etiferazioak kolesterola are gehiago hidrofoboa gordetzeko eta garraiatzeko.

Kolesterola hazten ari den animalia organismo bateko ehun guztietarako beharrezkoa da mintzak sintetizatzeko, eta organo batzuek (adibidez, guruin adrenalak eta sexu-guruinak) kolesterola hormona esteroideen aitzindari gisa erabiltzen dute (ondoren azalduko dugu). Kolesterola ere D bitaminaren aitzindaria da (ikus 10-20. Irudia, 1. bertsioa).
Kolesterola eta beste lipido batzuek plasma lipoproteinak eramaten dituzte
Kolesterola eta kolesterol esterrak, triacilglicerol eta fosfolipidoak bezala, ia uretan disolbaezinak dira. Hala ere, sintetizatu zuten ehunetik ehunetara biltegiratu edo kontsumituko diren ehunetara joan beharko dute. Odol-fluxuak darama odol-plasma lipoproteinak - eramaile proteinen berariazko konplexuen makromolekularrak (Apolipoproteins) Konbinatu hauetan fosfolipidoak, kolesterola, kolesterol esterrak eta triacilglicerolak daude.
Apolipoproteinak ("apo" lipidarik gabeko proteina bera aipatzen da) lipidoekin konbinatzen dira lipoproteinen partikulen hainbat zatiki osatzeko. Konplexu esferikoak erdian lipido hidrofoboekin eta aminoazido kate hidrofiloen azalean (Fig. 21-39, a). Lipidoen eta proteinen konbinazio desberdinak erabiliz, dentsitate desberdinetako partikulak eratzen dira - kromomikronetatik hasi eta dentsitate handiko lipoproteinak. Partikula hauek ultrazentrifugazio bidez bereiz daitezke (21-1. Taula) eta ikusmen behatu mikroskopia elektronikoa erabiliz (21-39. Irudia, b). Lipoproteinen frakzio bakoitzak funtzio zehatz bat betetzen du, sintesiaren lekua, lipidoen konposizioa eta apolipoproteinen edukiaren arabera zehazten dena. Gutxienez 10 apolipoproteina desberdin aurkitu dira giza odol-plasmaan (21-2 taula), tamainan aldatzen direnak, antigorputz zehatzak dituzten erreakzioak eta lipoproteinak klase desberdinetan banatzen diren ezaugarriak. Proteina-osagai hauek lipoproteinak ehun zehatzetara zuzentzen duten substantziak eta lipoproteinetan jarduten duten entzimak aktibatzen dituzte.
21-1 taula. Giza plasmako lipoproteinak
Oposizioa (masa frakzioa,%)
r = 513.000). LDLren partikula batek gutxi gorabehera 1.500 molekula kolesterolaren nukleoa dauka; nukleoaren inguruan 500 kolesterola, 800 fosfolipido molekula eta apoB-100 molekula bat daude. b - lipoproteinen lau klase, mikroskopio elektroniko batekin ikusgai (ezezko agerpenaren ondoren). Erlojuaren kontrako norabidea, ezkerreko goiko iruditik hasita: chilomikronak - 50 eta 200 nm arteko diametroa dutenak, PL O NP - 28 - 70 nm, HDL - 8 eta 11 nm bitartekoak, eta LDL - 20 eta 55 nm bitartekoak. Lipoproteinen propietateak taulan ematen dira. 21-2.

chylomicrons, aipatzen da Sec. 17, eraman elikagaien triacylglycerols hesteetatik beste ehunetara. Lipoproteinik handienak dira, dentsitate txikiena eta triacilglicerolen eduki erlatibo altuena dute (ikus 17-2. Irudia). Klomomroak heste txikiaren estalduraren zelula epitelialen ER sintetian sintetizatzen dira, eta ondoren sistema linfatikotik pasatu eta odolera sartzen dira ezkerreko subklaviako zainetatik. Chomomicron apolipoproteinak apoB-48 (lipoproteinen klase honetarako bakarra), apoE eta apoC-II (21-2 taula) daude. AroC-II-k lipoproteina lipasa aktibatzen du ehun adiposoen, bihotzaren, eskeletoko muskuluaren eta amaren guruineko amaren guruinetan. Horrela, klomomronek elikagaien gantz azidoak ehunetara transferitzen dituzte, eta bertan kontsumitu edo erregai gisa gordeko dira (Fig. 21-40). Chilomicron hondakinak (batez ere triacilgliceroletatik askatzen dira, baina oraindik kolesterola, apoE eta apoB-48) odoletik gibelera garraiatzen dira. Gibelean, errezeptoreak chilomicron-en hondakinetan jasotako apoE-rekin lotzen dira eta xurgapena endokitosiaren bidez bitartekatzen dute. Hepatozitoetan, hondakin horiek edukitzen duten kolesterola askatzen dute eta lisosometan suntsitzen dira.
21-2 taula. Giza plasma lipoproteina apolipoproteinak
Funtzioa (ezaguna bada)
L CAT aktibatzen du, eta ABC garraiatzailearekin elkarreragiten du
L CAT inhibitzen du
L CAT aktibatzen du, kolesterolaren garraioa / argia
LDL hartzaileari lotzen zaio
Chilomicrons, VLDL, HDL
Chilomicrons, VLDL, HDL
Chilomicrons, VLDL, HDL
VLDL eta chylomicron hondakinen garbiketa hasten da
Elikagaiak gaur egun erregai gisa erabil daitezkeen gantz-azido gehiago ditu, gibelean triazilglicerol bihurtzen dira, apolipoprotein espezifikoekin zatikia osatzen dutenak. oso dentsitate baxuko lipoproteinak (VLDL). Gibelean gehiegizko karbohidratoak triacilgliceroletara ere bihur daitezke eta VLDL gisa esportatu (Fig. 21-40, a).Triacilglicerolaz gain, VLDL frakzioak kolesterol eta kolesterol esteri kopuru jakin bat dauka, baita apoB-100, apoC-1, apoC-II, apoC III eta apoE (21-2 taula). Lipoproteinak hauek gibetik odoletik garraiatzen dira giharretara eta ehun adiposoetara, non, lipoproteina lipasa apo-C II aktibatu ondoren, gantz-azido askeak askatzen dira VLDL frakzioko triacilgliceroletatik. Adipozitoek gantz-azido askeak harrapatzen dituzte eta berriro triacilglicerol bihurtzen dira, zelula horietan gordetzen baitira lipidoen inklusioak (tantak), miokitoak, aitzitik, gantz-azidoak berehala oxidatzen dituzte energia sortzeko. VLDL hondakin gehienak zirkulaziotik hepatozitoak ezabatzen dira. Horien xurgapena, chomomikronen xurgapenaren antzekoa, hartzaileek bitartekatzen dute eta VEEL hondakinen apoE-ren presentziaren araberakoa da (21-2. Gehigarrian, apoE eta Alzheimer gaixotasunaren arteko erlazioa deskribatzen da).
Fig. 21-40. Lipoproteinak eta lipidoen garraioa, eta - lipidoak odoletik garraiatzen dira lipoproteinak direla eta, hainbat zatikitan konbinatzen dira funtzio desberdinak eta proteinen eta lipidoen konposizio desberdinak (21-1, 21-2 fitxa) eta zatiki horien dentsitateari dagokio. Elikagaien lipidoak ksilomikoetan biltzen dira, eta horietan dauden triazilglicerol gehienak lipoproteina lipasa askatzen dute kapiparretan muskulu-ehun adiposoetan. Klomomronoaren hondakinak (batez ere proteina eta kolesterola dauzkatenak) hepatozitoek harrapatzen dituzte. Lipido endogenoak eta gibeleko kolesterola adiposa eta giharretako ehunetara helarazten dira VLDL moduan. VLDL-ren lipidoak askatzeak (apolipoprotein batzuk galtzearekin batera) VLDLP LDL bihurtzen du pixkanaka, eta horrek kolesterola ehun extrahepatikoetara igortzen du edo gibelera itzultzen du. Gibelak VLDL, LDL eta chylomicrons-en aztarnak harrapatzen ditu errezeptorearen bitartekaritza endokitosian. Ehungabe gehiegizko kolesterola gibelera eramaten da LDL modura. Gibelean, kolesterolaren zati bat behazun gatz bihurtzen da. b - gosea hartu ondoren (ezkerrean) eta gantz-eduki handia duten eskuinean (eskuinean) janari odol-plasma laginak hartu. Elikagai koipetsuak jatean eratutako klomomronek plasma esnearekiko antzekotasuna ematen diote plasmari.

Triagliglizerolen galerarekin, VLDLren zati bat VLDL hondar bihurtzen da, tarteko dentsitateko lipoproteinak (VLDL) ere deitzen zaie. dentsitate baxuko lipoproteinak (LDL) (21-1. fitxa). LDL frakzioak, oso aberatsa da kolesterola eta kolesterola esterrak dituena eta apoB-100 ere badu, kolesterola transferitzen du hartzaile jakin batzuek beren plasma mintzetan aitob-100 aitortzen duten ehun extrahepatikoetara. Errezeptore hauek kolesterolaren eta kolesterol esterien kontsumoa bitartekatzen dute (jarraian deskribatzen den moduan).
21-2 gehigarria.ApoE aleloek alzheimerraren intzidentzia zehazten dute
Giza populazioan, apolipoproteina E. kodetzen duen genearen hiru aldaera ezagunak daude (apoE aleloen artean) APOEZ alela gizakien artean ohikoena da (% 78 inguru), APOE4 eta APOE2 aleloak 15 eta 7% dira, hurrenez hurren. APOE4 alela Alzheimerra duten pertsonen ezaugarria da bereziki, eta erlazio horrek probabilitate handia duen gaixotasuna gertatzea aurreikusten du. APOE4 heredatu duten pertsonek alzheimerra berandu garatzeko arrisku handia dute. APOE4-rako homozigotak dituzten pertsonek 16 aldiz gehiago dituzte gaixotasuna garatzeko; gaixo daudenen batez besteko adina 70 urte ingurukoa da. AROEZen bi ale heredatzen dituzten pertsonentzat, aitzitik, Alzheimerraren batez besteko adina 90 urtekoa da.
OraEOA eta Alzheimer gaixotasuna lotzeko oinarri molekularra oraindik ez da ezagutzen. Gainera, oraindik ez dago argi nola apoE4-k amiloide kableen hazkundean eragina izan dezakeen, itxuraz Alzheimer gaixotasunaren kausa nagusia (ikus 4-31. Irudia, 1). Hipotesi neuronen zitoskeletoaren egitura egonkortzeko apoE-ren balizko balorean oinarritzen da. ApoE2 eta apoEZ proteinak neuronaren mikrotubuluekin lotutako proteina batzuekin lotzen dira, eta apoE4 ez da lotzen. Horrek neuronen heriotza azkartu dezake. Mekanismo hori edozein izan daitekeen arren, behaketa horiek apolipoproteinen funtzio biologikoak gure ulermena zabaltzeko itxaropena ematen dute.
Laugarren lipoproteinak - Dentsitate handiko lipoproteinak (HDL), Zatiketa hori gibelean eta heste txikian sortzen da, proteina ugari duten partikula txikien kolesterola nahiko gutxi dutenak eta kolesterola esterorik gabe (21-40. irudia). HDL frakzioak apoA-I, apoC-I, apoC-II eta beste apolipoproteinak ditu (21-2 taula), baita lecitina-kolesterola-aziltransferasa (LC AT), lezitinatik (fosfatidilkolina) eta kolesterotik (kolesterol esterrak) sortzen direla katalizatzen duena (21-41 irudia). LT-k sortutako HDL partikulen gainazalean L CAT-k kolesterol kolesterola eta fosfatidilkolina eta VLDL hondarrak bihurtzen ditu, nukleoa eratzen hasten direnak, sortu berri diren HDL partikula diskidoak HDL partikula esferiko helduak bihurtuz. Kolesterola daukan lipoproteina gibelera itzultzen da eta bertan kolesterola "deskargatzen" da, kolesterol hori zati bat gatz gatz bihurtzen da.
Fig. 21-41. Lezitina-kolesterola-aziltransferasaren bidez katalizatutako erreakzioa (L CAT). Entzima hau HDL partikulen azalean dago eta apoA-1 aktibatuta dago (HDL frakzioaren osagaia). Kolesterola-esterrak sortu berri diren HDL partikulen barruan pilatzen dira eta horiek HDL helduak bihurtzen dira.

HDL gibelean xurgatu daiteke endokitosiaren bidez, baina gutxienez HDL kolesterola beste ehun batzuetara igortzen da beste mekanismo batzuen bidez. HDL partikulek SR - BI hartzailearen proteinekin lotu daitezke gibeleko zelulen plasmako mintzetan eta ehun esteroideogenikoetan, guruin adrenaletan. Hartzaile hauek ez dute endokitosia bitartekatzen, baizik eta kolesterola eta HDL frakzioaren beste lipidoak zeluletara transferitzea. HDL zatiketa "agortua" berriro odolera sartzen da eta bertan, kromomikronen eta VLDL hondakinen lipidoen zati berriak sartzen dira. HDL berberak ehun extrahepatikoetan gordetako kolesterola ere eraman dezake eta gibelean transferitu kolesterolaren alderantzizko garraioa (21-40. Irudia). Alderantzizko garraio aldaeraren batean, kolesterolaren zeluletan SRL-BI hartzaileekin SR-BI hartzaileen elkarrekintzak kolesterolaren zelularen hedapen pasiboa hasten du zelulen gainazaletik HDL partikuletara eta, ondoren, kolesterola gibelera transferitzen du. Kolesterolaren zelula aberatseko alderantzizko garraioaren beste aldaera batean, HDL zatikatu ondoren, apoA-I garraiatzaile aktiboarekin interagitzen da ABC proteinarekin. ApoA-I (eta ustez HDL) endokitosiaren bidez xurgatzen da, gero berriro jariatzen da, kolesterola kargatuta, gibelean garraiatzen dena.
ABC1 proteina droga askoren eramaileen familia ugarikoa da. Eramaile horiek ABC garraiatzaileak deitzen dira batzuetan. Izan ere, guztiek ATP lotesisten kaseteak dituzte (ATP - lotzen diren kaseteak), transmembranen sei domeinu ere badituzte sei transmembrana helizeekin (ikus kap. . 11, v. 1). Proteina horiek aktiboki ioiak, aminoazidoak, bitaminak, hormona esteroideak eta behazun-gatzak transferitzen dituzte plasma mintzen bidez. Garraiolari familia horren beste ordezkari bat CFTR proteina da, fibrosi kistikoa kaltetuta baitago (ikus 11-3. Eranskina, 1. paragrafoa).
Kolesterola esterrak zelulan sartzen dira endokitosiaren bidez hartutako bitartekoen bidez
Odol-fluxuan dagoen LDL partikula bakoitzak apoB-100 dauka, gainazal hartzaile espezifikoko proteinek ezagutzen dutena -LDL hartzaileak kolesterola harrapatu behar duten zelulen mintzetan. LDL hartzaileak LDL hartzaileari endokitosia eragiten dio. Horregatik, LDL eta haren hartzailea endosomaren barnean dagoen zelulara mugitzen dira (Fig. 21-42). Endosoma azkenean lisosomarekin nahasten da, eta honek kolesterola esterrak hidrolizatzen dituzten entzimak ditu, kolesterola eta gantz azidoak zitosolera askatuz. LDL-ren ApoB-100-ek ere, zitosoletan jariatzen diren aminoazidoak eratzeko, baina LDL hartzaileak degradazioa ekiditen du eta zelularen gainazalera itzultzen da berriro LDL hartzean parte hartzeko. ApoB-100 VLDL-n ere badago, baina hartzaileari lotzen zaion domeinua ez da LDL hartzaileari lotzeko gai; VLDLP LDL bihurtzeak errezeptorearen lotura duen domeinua apoB-100 bihurtzea lortzen du. Odoleko kolesterola garraiatzeko bide hori eta xede-ehunetan hartutako endokitosia bitartekotutako endokitosia Michael Brown eta Joseph Goldstein-ek aztertu dute.
Michael Brown eta Joseph Goldstein

Fig. 21-42. Kolesterolaren harrapaketa endokitosiaren bidez hartutako indarrarekin.

Horrela, zeluletara sartzen den kolesterola mintzetara sartu edo ACAT (21-38 irudia) berriderifikatu daiteke lipido tanta zitosoletan gordetzeko. Odoleko LDL zatikian kolesterol nahikoa eskuragarri dagoenean, gehiegizko kolesterola zelulen gehiegizko metaketa saihesten da, bere sintesia tasa murriztuz.
LDL hartzailea apoE-rekin ere lotzen da eta gibelarekiko chylomicrons eta VLDL hondarrak hartzerakoan paper garrantzitsua du. Hala ere, LDL hartzaileak eskuragarri ez badaude (adibidez, LDL hartzailearen falta duen sagu-tentsioan), VLDL hondarrak eta klomomronoak gibelean xurgatzen dira, nahiz eta LDL xurgatu ez. Honek, VLDL eta chylomicron hondakinen endokitosi bitartekariaren hartzaileentzako erreserba sistema laguntzaile baten presentzia adierazten du. Erreserbaren hartzaileetako bat LRP proteina da (lipoproteinen errezeptorearekin lotutako proteina), lipoproteinen hartzaileekin erlazionatuta dagoena, apoE eta beste ligando batzuekin lotzen dena.
Kolesterolaren biosintesia erregulatzeko hainbat maila
Kolesterolaren sintesia prozesu konplexua eta energetikoki garestia da, eta, beraz, argi dago gorputzak onuragarria duela kolesterolaren biosintesia erregulatzeko mekanismo bat izatea, eta horrek elikadurarekin batera datorren kopurua berregiten du. Ugaztunetan, kolesterol-produkzioa zelulen arteko kontzentrazioaren bidez arautzen da
kolesterola eta hormonak glukagona eta intsulina. HMG - CoA bihurtzeko fasea mevalonatera (Fig. 21-34) kolesterolaren eraketa metabolikoaren bide metabolikoan abiadura mugatzen da (erregulazio puntu nagusia). Erreakzio hau HMG - CoA reductase bidez katalizatzen da. Kolesterolaren mailen aldaketen aurrean erregulazioa HMG - CoA reductase kodetzen duen gene baten transkripzio erregulazio sistema dotore baten bidez egiten da. Gene hau, kolesterolaren eta gantz-azido insaturatuen xurgapenean eta sintesian parte hartzen duten beste 20 gene baino gehiagorekin batera, proteina eratzen duten esteroreregulatzaileekin elkarreragiten duten proteina familia txiki batek kontrolatzen du (SREBP, esterolaren elementu erregulatzaileen loturak). . Sintesi ondoren, proteina hauek erretululu endoplasmikoan sartzen dira. SREBP domeinu aminoazterreko soluzio bakarrak transkripzio-aktibatzaile gisa funtzionatzen du Ch-en deskribatutako mekanismoak erabiliz. 28 (3. v.). Hala ere, domeinu honek ez du nukleorako sarbiderik eta ezin du genearen aktibazioan parte hartu SREBP molekulan egon den bitartean. HMG genearen transkripzioa aktibatzeko - CoA reductasa eta beste geneak, transkripzio aktiboaren domeinua gainerako SREBP bereizten da zatiketa proteolitikoaren bidez. Kolesterola altua denean, SREBP proteinak ez-aktiboak dira, ER batean finkatuta SCAP (SREBP - cleavage activating protein) beste konplexu batean (Fig. 21-43). SCAP da kolesterola eta beste hainbat esterol lotzen dituena, esterol sentsore gisa jarduten duena. Esterol maila handia denean, SCAP - SREBP konplexuak seguruenik beste proteina batekin interakzionatzen du, eta horrek konplexutasun osoa mantentzen du ER-an. Zelularen esterolen maila jaisten denean, SCAPen konformazio aldaketak atxikipen jarduera galtzea dakar eta SCAP - SREBP konplexua besikularen barnean Golgi konplexura migratzen da. Golgi konplexuan, SREBP proteinak bi proteasa desberdin bikoizten dira, bigarren giltza amino-terminal domeinua zitosolera askatuz. Domeinu hau nukleora mugitzen da eta xede geneen transkripzioa aktibatzen du. SREBP proteina domeinu amino-terminalak erdibide laburra du eta proteasomek azkar degradatzen dute (ikus 27-48. Irudia, 3. irudia). Esterolaren maila nahikoa igotzen denean, SR EBP proteina domeinuen askapen proteolitikoa amino terminoarekin blokeatu egiten da berriro, eta lehendik dauden domeinu aktiboen proteasomaren degradazioak xede geneak azkar ixtea eragiten du.
Fig. 21-43. SR EBP aktibatzea. SREB P proteinak esterol bidez erregulatutako elementu batekin (kolore berdea) elkarreragiten dutenak, sintesia egin eta berehala, ERan sartzen dira, S CAP (kolore gorria) konplexua osatzen dutenak. (N eta C proteinen amina eta karboxilen muturrak adierazten dira.) S-CAP lotutako egoeran, SRE BP proteinak ez daude. Esterolaren maila jaisten denean, SR EBP-S CAP konplexua Golgi konplexura migratzen da, eta SR EBP proteinak sekuentzialki bi proteasak bereizten dituzte. Askatutako aminoazido terminal SR EBP proteinaren domeinua nukleora migratzen da, eta bertan esterolek erregulatutako geneen transkripzioa aktibatzen du.

Kolesterolaren sintesia beste hainbat mekanismoren bidez ere arautzen da (21-44. Irudia). Kontrol hormonalak NM G-CoA reductasearen aldaketa kobalentearen bidez egiten du eragina. Entzima hau fosforilatu (aktibo) eta desfosforilatu (aktibo) dauden formetan existitzen da. Glucagonak entzimaren fosforilazioa (inaktibazioa) estimulatzen du eta intsulinak desfosforilazioa bultzatzen du, entzima aktibatuz eta kolesterolaren sintesia hobetuz. Kolesterola zelulen arteko kontzentrazio altuek ASAT aktibatzen dute, eta horrek kolesterola metaketaren esterifikazioa areagotzen du. Azkenik, kolesterol zelular maila altuek LDL hartzailea kodetzen duen gene baten transkripzioa inhibitzen dute, hartzaile horren produkzioa murriztuz eta, beraz, kolesterola odoletik hartuz.
Fig. 21-44. Kolesterolaren maila erregulatzeak elikagaien kolesterolaren sintesia eta xurgapenaren arteko oreka ematen du. Glucagonak NM G-CoA reductasearen fosforilazioa (desaktibazioa) errazten du, intsulinak defosforilazioa (aktibazioa) sustatzen du. X - identifikatu gabeko kolesterol metabolitoak NM G -CoA reductasearen proteolisia estimulatzen dutenak.

Erregulatu gabeko kolesterolak gaixotasun larriak ekar ditzake gizakietan. Elikagaietatik lortutako kolesterolaren eta kolesterol sintetikoen kopuru osoa mintz-muntaiarako beharrezko zenbatekoa gainditzen denean, gatz-gatz eta esteroideen sintesia, odol-hodietan (plaka aterosklerotikoak) kolesterolaren metaketa patologikoak ager daitezke, eta hori blokeatu egiten da (aterosklerosia). Herrialde industrializatuetan bihotz-gutxiegitasuna da arteria koronarioak heriotzaren kausa nagusia dela eta. Aterosklerosiaren garapena odol kolesterol maila altua eta batez ere LDL frakzioarekin transferitutako kolesterol maila altua eta HDL odol maila altuak, aitzitik, onuragarria izaten dute odol hodien egoeran.
Hiperkolesterolemia hereditarioarekin (akats genetikoa), odol kolesterolaren maila oso altua da - aterosklerosi larria garatzen da pertsona haurtzaroan. LDL hartzaile akastuna dela eta, ez da nahikoa hartzailearen bitartekaritza hartzailea LDL kolesterola. Ondorioz, kolesterola ez da odoletik kentzen, pilatzen da eta plaka aterosklerotikoak eratzen laguntzen du. Kolesterola endogenoaren sintesia odolean gehiegizko kolesterola izan arren, zelulaz kanpoko kolesterola ezin da zeluletara sartu zelulen arteko sintesia erregulatzeko (Fig. 21-44).Hiperkolesterolemia hereditarioa duten eta gaixoen kolesterolarekin lotutako beste gaixotasun batzuk tratatzeko, estatina klaseak erabiltzen dira. Horietako batzuk iturri naturaletatik lortzen dira, eta beste batzuk, berriz, industria farmazeutikoak sintetizatzen ditu. Estatinak mevalonatoaren antzekoak dira (21-3 gehigarria) eta NMS-CoA reductasearen inhibitzaile lehiakorrak dira.
21-3 gehigarria. MEDICINE. Lipidoen hipotesia eta estatinak sortzea
Bihotzeko gaixotasun koronarioak (CHD) heriotza-kausa nagusia da herrialde garatuetan. Odola bihotzera daraman arteria koronarioen murrizketa plaka aterosklerotiko izeneko gantz gordailuak eratzearen ondorioz gertatzen da; plaka hauek kolesterola, fibrilar proteinak, kaltzioa, plaketetako koadroak eta zelula zatiak dituzte. Mendean XX. Eztabaida aktiboa egon zen arteriaren obstrukzioaren (aterosklerosi) eta odol kolesterolaren arteko harremanari buruz. Norabide horretan eztabaida eta ikerketa aktiboak kolesterola jaisten duten sendagai eraginkorrak sortzea eragin dute.
1913an, N. N. Anichkov-ek, errusiar zientzialari ezagunak eta patologia esperimentalaren arloan aditua denak, lan bat argitaratu zuen, bertan erakutsi zuen kolesterol ugari duten jakiekin elikatutako untxiek kalteak sortzen dituztela adineko pertsonen ontzietan plaka aterosklerotikoen antza duten odol hodietan. Anichkov-ek hainbat hamarkadatan egin zuen ikerketa eta emaitzak Mendebaldeko aldizkari ezagunetan argitaratu zituen. Zoritxarrez, bere datuak ez ziren gizakien aterosklerosia garatzeko eredu baten oinarria bihurtu, garai hartan hipotesia nagusitzen ari zela gaixotasun hori zahartzearen emaitza naturala dela eta ezin dela prebentzioa izan. Hala ere, frogak pilarearen kolesterol serumaren eta aterosklerosiaren (hipotesi lipidikoen) eta 1960ko hamarkadan garatzen ari ziren harremanak pilatzen joan ziren. Zenbait ikertzailek adierazi dute esplizituki gaixotasun hau sendagaiekin tratatu daitekeela. Hala ere, aurkako ikuspuntua egon zen 1984an AEBetako Osasun Institutu Nazionalak (Koronarioen Prebentzio Proba) egindako kolesterol-eginkizunaren ikerketa zabal baten emaitzak argitaratu ziren arte. Miokardioko infartuen maiztasuna eta odol kolesterolaren gutxitzeak eragindako kolpeak estatistikoki nabarmen murriztu ziren. Azterketa honetan, kolesterola, behazun azidoak lotzen dituen erretxina trukatzeko errolina kolesterola jaisteko erabili zen. Emaitzek sendagai terapeutiko indartsuagoak bilatzea bultzatu dute. Esan behar dut mundu zientifikoan lipideen hipotesiaren balioei buruzko zalantzak erabat desagertu zirela estatinen etorrerarekin bakarrik 80ko hamarkadaren amaieran - 1990eko hamarkadaren hasieran.
Lehen estatina Akira Endo-k aurkitu zuen Tokioko Sankyo-n. Endo-k 1976an argitaratu zuen bere lana, nahiz eta hainbat urtez kolesterolen metabolismoaren arazoa jorratu zuen. 1971n, kolesterolaren sintesiaren inhibitzaileak garai hartan aztertutako antibiotikoen onddoen ekoizleak ere egon zitezkeela proposatu zuen. Hainbat urtetan lan intentsiboan, hainbat perretxiko 6.000 kultura baino gehiago aztertu zituen, emaitza positiboa lortu zuen arte. Emaitzen den konposatuari compactin deitzen zitzaion. Substantzia horrek kolesterola txikiagotu zuen txakurretan eta tximinoetan. Ikerketa horiek Texas Brown-eko hego-mendebaldeko Medikuntza Eskolako Michael Brown eta Joseph Goldstein-en arreta jaso zuten. Brownek eta Goldstein-ek, Endorekin batera, ikerketa bateratua hasi zuten eta bere datuak berretsi zituzten. Lehen entsegu klinikoen arrakasta handiek farmazia konpainiek drogen berri horien garapenean parte hartu zuten. Merck-en, Alfred Alberts-ek eta Roy Wagelos-ek zuzendutako talde batek onddo kulturen proiekzio berria jarri zuen abian eta, guztira, 18 kulturatan aztertuta, beste droga aktibo bat aurkitu zuten. Substantzia berria lovastatina deritzo. Hala ere, aldi berean, asko uste zen compactin dosi altuak txakurrei emateak minbizia garatzea eta estatina berriak bilatzea 1980ko hamarkadan. eten egin da. Hala ere, garai hartan, familiaren hiperkolesterolemia gaixoak tratatzeko estatinak erabiltzearen abantailak jada baziren. Nazioarteko adituekin eta Elikagaien eta Droga Administrazioarekin (FDA, AEB) kontsultak egin ondoren, Merck lovastatina garatzen hasi zen. Hurrengo bi hamarkadetan egindako azterlan ugariek ez dute agerian lovastatinaren eragin kartzinogenoa eta ondoren agertu diren drogen sorrera berria.
Fig. 1. Estatinak NM G-CoA reductasearen inhibitzaileak dira. NM G -CoA reductasearen ekintza inhibitzen duten mevalonatoaren eta lau farmazia produktuen (estatinak) egitura konparatzea.

estatina HMG - CoA - reductase-ren ekintza inhibitu, mevalonatoaren egitura imitatuz eta, horrela, kolesterolaren sintesia blokeatu. LDL hartzailearen genearen kopia bateko akats batek eragindako hiperkolesterolemia duten gaixoetan, lovastatina hartzerakoan, kolesterol maila% 30 murrizten da. Botika are eraginkorragoa da behazun-azidoak lotzen dituzten erretxina bereziekin konbinatuz eta hesteetatik alderantzizko xurgapena ekiditen dutenean.
Gaur egun, estatinak gehien erabiltzen dira odol plasmako kolesterola jaisteko. Edozein botika hartzerakoan, nahi ez diren bigarren mailako efektuei buruzko galdera sortzen da. Hala ere, estatinen kasuan, bigarren mailako efektu asko positiboak dira. Droga hauek odol-fluxua estimulatu dezakete, lehendik zeuden plaka aterosklerotikoak konpondu (odol-hodietako hormetatik ez hausteko eta odol-fluxua oztopatzeko), plaketen agregazioa inhibitu eta odol hodietako hormetan hanturazko prozesuak ahultzen dituzte. Lehen aldiz estatinak hartzen dituzten pazienteetan, efektu horiek kolesterolaren maila jaisten hasi aurretik ere agertzen dira eta isoprenoideen sintesia inhibitzearekin lotzen dira. Jakina, estatinen alde guztietan ez da onuragarria. Zenbait gaixoetan (normalean kolesterola jaisten duten beste droga batzuekin batera estatinak hartzen dituztenen artean), muskulu mina eta gihar ahultasuna gerta daitezke eta, batzuetan, nahiko modu gogorrean. Estatinen bigarren mailako efektu nahiko ugariak ere erregistratzen dira, zorionez oso gutxitan gertatzen direnak. Gaixoen gehienetan, estatinak hartzeak gaixotasun kardiobaskularrak garatzea galarazi dezake. Beste edozein botika bezala, estatinak zure medikuak gomendatzen duen moduan bakarrik erabili behar dira.

HDL kolesterolaren herentziarik gabe, kolesterol maila oso baxuak dira, Tangerreko gaixotasunarekin kolesterola ia ez dago zehaztua. Bi gaixotasun genetikoak ABC1 proteinaren mutazioen ondorioz sortzen dira. HDLik gabeko kolesterolaren frakzioak ezin du kolesterola ABC1 defizita duten zeluletatik harrapatu eta kolesterola agortzen duten zelulak odoletik azkar kendu eta suntsitu egiten dira. HDL eta Tangier gaixotasunaren oinordetza eza oso bakanak dira (Tanger gaixotasuna duten 100 familia baino gutxiago ezagutzen dira mundu osoan), baina gaixotasun horiek ABC1 proteinaren HDL plasma maila erregulatzeko papera erakusten dute. Plasma HDL maila baxuak arteria koronarioen kalte tasa handiarekin erlazionatzen direnez, ABC1 proteina xede erabilgarria izan daiteke HDL maila erregulatzeko diseinatutako sendagaientzat. ■
Hormona esteroideak kolesterolaren alboko katea eta haren oxidazioa zatituz osatzen dira.
Pertsona batek bere hormona esteroide guztiak kolesterolagatik jasotzen ditu (21-45. Irudia). Hormona esteroideen bi klase sintetizatzen dira adrenaleko kortexean: mineralkortikoidy,ioi inorganikoen xurgapena erregulatzen dutenak (Na +, C l - eta HC O) 3 -) giltzurrunetan, eta glucocorticoids, glukoneogenesia erregulatzen eta hanturazko erantzuna murrizten laguntzen dutenak. Sexu-hormonak gizonen eta emakumeen ugalketa-zeluletan eta plazentan sortzen dira. Horien artean progesterone, emakumezkoen ugalketa zikloa erregulatzen duena, androgens (adibidez, testosterona) eta estrogens (estradiol), gizonezkoen eta emakumezkoen bigarren mailako ezaugarri sexualen garapenean eragina dutenak, hurrenez hurren. Hormona esteroideek oso kontzentrazio baxuetan dute eragina eta, beraz, kantitate nahiko txikietan sintetizatzen dira. Hesietako gatzekin konparatuta, hormona esteroideak ekoizteko kolesterol gutxi kontsumitzen da.

Fig. 21-45. Hormona esteroide batzuk kolesteroletik eratzen dira. Konposatu horietako batzuen egiturak irudian ageri dira. 10-19, 1. v.

Hormona esteroideen sintesiak karbono C-17 D-eraztunaren "alboko katean" karbono atomo ugari edo guztiak kentzea eskatzen du. Bigarren mailako kate kentzea ehun steroidogenikoen mitokondrian gertatzen da. Kentzeko prozesua alboko katearen bi karbono atomoen hidroxilazioan datza (C-20 eta C-22), eta gero haien arteko lotura zatikatzea (Fig. 21-46). Hainbat hormonen eraketa oxigeno atomoak ere sartzen dira. Esteroideen biosintesi zehar hidroxilazio eta oxidazio erreakzio guztiak funtzio mistoko oxidasesek katalizatzen dituzte (21-1 gehigarria) NА D PH, O erabiltzen dutenak 2 eta mitokondriako P-450 zitokromoa.
Fig. 21-46. Alboko katea zatitzea hormona esteroideen sintesian. Aldameneko karbono atomoak oxidatzen dituen funtzio mistoa duen oxidase sistema honetan, P-450 zitokromoak elektroien eramaile gisa jokatzen du. Prozesuan ere parte hartzen dute proteinak, adrenodoxina eta adrenodoxina murriztasa garraiatzeko. Alboko kateen zatitze sistema hori adrenaleko kortexaren mitokondrian aurkitu zen, eta bertan esteroideen produkzio aktiboa gertatzen da. Pregnenolona beste hormona esteroide guztien aitzindaria da (Fig. 21-45).

Kolesterolaren biosintesi bitartekariak beste bide metaboliko askotan parte hartzen dute.
Kolesterolaren biosintesi bitarteko gisa duen eginkizunaz gain, isopentenil pirofosfatoa aktibatutako aitzindaria da. Hainbat funtzio biologiko betetzen dituzten biomolekula ugari sintetizatzeko (21-47. Irudia). Horien artean, A, E eta K bitaminak, karoteno eta klorofilen fitol katea bezalako landare-pigmentuak, kautxu naturala, funtsezko olio ugari (adibidez, limoi olioaren oinarri usaingarria, eukaliptoa, muskia), metamorfosia erregulatzen duen hormona gazteen intsulak, dolikolak. Lipido disolbagarriak eraiki polysaccharides, ubiquinone eta plastoquinone sintesi konplexu gisa - elektroi eramaileak mitokondrian eta kloroplastetan. Molekula horiek guztiak egitura isoprenoideak dira. 20.000 isoprenoide baino gehiago aurkitu dira naturan, eta ehunka berri berri ematen dira urtero.
Fig. 21-47. Isoprenoideen biosintesiaren irudi orokorra. Hemen aurkezten diren azken produktuen gehienen egiturak kapituluan ematen dira. 10 (v. 1).

Prenilazioa (isoprenoide baten atxikimendu kobalentea, ikus 27-35. Irudia) mekanismo arrunta da, proteinen bidez aingura daitekeen zelula ugaztunen mintzetan (ikus 11-14 irudia). Proteina batzuetan, lotutako lipidoa 15 karbono-farnesil talde batek irudikatzen du, beste batzuetan 20 karbonoko geranil geranilo taldea da. Bi lipido mota hauek entzima ezberdinak lotzen dituzte. Litekeena da prenilazio erreakzioek proteinak mintz desberdinetara zuzentzea lipidoa atxikita dagoen arabera. Proteina erretilatzea isoprene deribatuentzat beste eginkizun garrantzitsua da. Kolesterol bide metabolikoaren parte-hartzaileak.
21.4 atalaren laburpena Kolesterol, esteroide eta isoprenoideen biosintesia
■ Kolesterola acetil-CoA-tik erreakzio sekuentzia konplexuan eratzen da, hala nola, β-hydroxy-β-metylglutaryl-CoA, mevalonate, isoprene dimetilalil pirofosfatoa eta isopentenil pirofosfatoa aktibatuta. Unitate isoprenoen kondentsazioak squalene ez ziklikoa ematen du, zirkulatzen duena kondentsatutako eraztun sistema eta esteroideen alboko katea osatzeko.
■ Kolesterolaren sintesia hormonalen kontrolpean dago eta, gainera, kolesterola zelulen arteko kontzentrazioak handituz inhibitzen da, aldaketa kobalentearen eta transkripzioaren erregulazioaren bidez gertatzen baita.
■ Kolesterola eta kolesterola esterrak odolak eramaten ditu plasma lipoprotein gisa. VLDL frakzioak kolesterola, kolesterola esterrak eta triacilglicerolak gibetik transferitzen ditu beste ehunetara, non triacilglicerolak lipoproteinen lipasa bidez ebakitzen diren eta VLDL LDL bihurtzen den. Kolesterolaren eta kolesterolaren esterietan aberastutako LDL zatiketa endozitosiaren bidez zeharka harrapatzen da, eta LDLko B-100 apolipoproteinak, berriz, plasmako mintz hartzaileek aitortzen dute. HDLak kolesterola odoletik kendu, gibelera transferituz. Kolesterolaren metabolismoan nutrizio baldintzek edo akats genetikoek aterosklerosia eta miokardio infartua ekar dezakete.
■ Esteroideen hormonak (glukokortikoideak, mineralokortikoideak eta sexu-hormonak) kolesterotik sortzen dira alboko katea aldatuz eta eraztunen esteroideen sisteman oxigeno atomoak sartuz. Beste konposatu isoprenoide asko mevalonatatik sortzen dira isopentenil pirofosfatoaren eta dimetilalil pirofosfatoaren kondentsazioarekin.
■ Zenbait proteinen hartzidura zelulak mintzekin lotzeko guneetara zuzentzen da eta oso garrantzitsua da beren jarduera biologikorako.
48. galdera. Gantz-azido altuen metabolismoaren erregulazioa (β-oxidazioa eta biosintesia). Malonil CoAren sintesia. Acetyl CoA carboxylase, bere jardueraren erregulazioa. Mitokondriaren barneko mintzetik azido Co-a garraiatzea.
nagusia
fenilalanina kantitatea kontsumitzen da
2 eratara:
pizten da
urtxintxetan
bihurtutako
tirosinan.
eraldaketa
fenilalanina tirozina nagusiki
gehiegizkoa kentzeko beharrezkoa
fenilalanina, kontzentrazio altuak baitira
Zelulentzat toxikoa da. eraketa
tirosina ez du axola
aminoazido honen gabezia geroztik
zeluletan ia ez da gertatzen.
lehen
fenilalaninaren metabolismoa hasten da
hidroxilazioarekin (9-29. irudia), alegia
tirosina sortuz.
Erreakzio hori zehatz batek katalizatzen du
monooxy-nase - fenilalanine hydra (zsilasa,
koproduktore gisa balio duena
tetrahidrobiopterina (N4BP).
Entzimaren jarduera ere horren araberakoa da
Fe2-ren presentzia.
The
gibela batez ere bizkortutako mobilizazioa da
glukogenoa (ikus 7. atala). Hala ere, izakinak
Gibelean glukogenoa agortu egiten da
18-24 ordu barau. Iturri nagusia
glukosa izakinak agortu ahala
glukogenoa glukoneogenesi bihurtzen da,
bidez azeleratzen hasten dena
Fig.
11-29. Aldaketa metaboliko garrantzitsuak
energia xurgatzailea aldatzean
egoera postabsorbente. CT
- zetona gorputzak, FA - gantz azidoak.
4-6 h
azken bazkariaren ostean. substratu
glizerola glukosa-sintesia egiteko erabiltzen da,
aminoazidoak eta laktatoa. Altua
glukagonoaren kontzentrazioaren sintesi tasa
gantz-azidoak murriztu egin dira
fosforilazioa eta aktibazioa
azetilen CoA karboxilasa eta tasa
p-oxidazioa handitzen da. Aldi berean
gantz hornidura handitu gibelean
garraiatzen diren azidoak
gantz biltegietatik Azetil-CoA eratu da
gantz-azidoen oxidazioan erabiltzen da
gibelean zetona-gorputzen sintesia egiteko.
The
ehun adiposoak kontzentrazio handituz
glukagonoak sintesi tasa murriztua
TAG eta lipolisia estimulatzen dira. estimulazioa
lipolisia - aktibazio emaitza
hormona-sentikorra TAG lipasa
adipozitoak glukagonaren eraginpean.
Gantz-azidoak garrantzitsuak dira
energia iturriak gibelean, giharretan eta
ehun adiposa.
Hori dela
beraz, postabsorzio aldian
odol glukosa kontzentrazioa mantentzen da
80-100 mg / dl-ko mailan, eta gantz-maila
azidoak eta zetona gorputzak ugaritzen dira.
azukre
diabetesa gertatzen den gaixotasuna da
absolutua edo erlatiboa dela eta
intsulina gabezia.
A.
Azukrearen forma kliniko nagusiak
diabetes
Arabera
Mundu Erakundea
osasun-diabetesa
ezberdintasunen arabera sailkatuta
faktore genetikoak eta klinikoak
bi forma nagusi: diabetesa
I Mota - intsulinaren menpekoa (IDDM), eta diabetesa
II mota - intsulina ez den independentea (NIDDM).
erregulazio
zhk-ren sintesia. entzima erregulatzailea
lcd - acetyl CoA carboxylase-ren sintesia.
Entzima hori hainbat arautzen da
moduak.
Aktibazioa / Disoziazioa
entzima azpitaldeko konplexuak. The
acetyl CoA carboxylase forma inaktiboa
konplexuak bereizten ditu,
horietako bakoitza 4 azpitaldez osatuta dago.
Entzimaren aktibatzailea citrato da. Estimulatzen du
konplexuen konbinazioa, ondorioz
eta horren bidez, entzimaren jarduera handitzen da
. Inhibitzaile-palmitoyl-CoA. Deitzen zuen
konplexua disoziazioa eta jaitsiera
entzimaren jarduera.
Fosforilazioa / Desfosforilazioa
acetil CoA carboxylase. The
postabsorzio egoera edo
lan fisikoa glukgonizatua
adrenalina adenilato ziklozearen bidez
sistema prokinasa A eta aktibatuta dago
azpiunitatearen fosforilazioa bultzatzea
acetil CoA carboxylase. phosphorylated
entzima aktiboa da eta gantz sintesia
azidoak gelditzen dira.
absorptive The
period intsulinak fosfatasa aktibatzen du,
eta azetil-CoA karboxilasa sartzen da
egoera desfosforilatua gero
zitratuaren eraginpean gertatzen da
entzimaren protomeroen polimerizazioa, eta
aktibo bihurtzen da. Aktibazioaz gain
entzima citratoak beste bat egiten du
funtzioa LCD sintesian. absorptive The
gibeleko zelulen mitokondrian periodoa
citrato pilatzen da, eta horretan
azidoaren hondakina hona eramaten da
cytosol.
erregulazio
β-oxidazio tasak.
Oxid-oxidazio-bide metabolikoa,
lotura estua du CPEren eta orokorraren lanarekin
katabolismo moduak. Hori dela eta, bere abiadura
zelularen beharra araututa
energia i.e. ATP / ADP eta NADH / NAD ratioak direla eta, baita erreakzio tasa CPE eta
katabolismoaren bide arrunta. abiadura
Ehunen β-oxidazioa erabilgarritasunaren araberakoa da
substratua, hau da.
gantz kantitatearen gainean
mitokondrioetan sartzen diren azidoak.
Gantz azido askeen kontzentrazioa
odolean aktibazioa igotzen da
lipolisia ehun adiposoan barau garaian
glukagonaren eraginpean eta fisikoan zehar
adrenalinaren eraginpean lan egitea. Hauetan
gantz azido bihurtzen dira
energia iturri nagusi
giharretarako eta gibelerako, ondorioz
β-oxidazioak NADH eta azetil-CoA inhibitzaileek osatzen dituzte
piruvato deshidrogenasa konplexua.
Piruvatoaren eraketa
glukosa etatik azetil-CoA moteldu egiten da.
Bitarteko metabolitoak metatzen dira
glikolisia eta, bereziki, glukosa-6-fosfatoa.
Glukosa-6-fosfatoak hexokinasa inhibitzen du
eta, beraz, baztertzen du
glukosa prozesuan erabiltzea
glycolysis. Horrela, nagusi
lcd erabiltzea iturri nagusi gisa
energia muskulu-ehunetan eta gibelean
nerbio ehunetarako glukosa aurrezten du
globulu gorriak.
Β-oxidazio tasa ere bai
entzimaren jardueraren araberakoa da
karnitina akiltransferasak I.
Gibelean entzima hori inhibitzen da.
malonil CoA, eratutako substantzia
lcd-ren biosintesiarekin. Xurgapen-aldian
glicolisia gibelean aktibatzen da eta
azetil-CoAren eraketa handitzen da
piruvatoa. Lehen sintesia erreakzioa
lcd bihurketa acetyl-CoA malonil-CoA.
Malonil-CoA-k lcd-ren β-oxidazioa inhibitzen du,
sintesia egiteko erabil daitekeena
gantz.
eraketa
malonil-CoA azetil-CoA-erregulatzaileetatik
biosintesia lcd erreakzioa. Lehenengo erreakzioa
sintesia lcd bihurketa azetil-CoA-ra
malonil CoA. Entzima katalitikoa
erreakzio hau (acetil Coa carboxylase),
ligasaren klasekoak dira. Berak dauka
kobalenteki loturiko biotina. Lehenengoan
erreakzio kobalenteen faseak
Energia dela eta biotina lotzen du
ATP, 2. fasean COO transferentzia
azetil-CoA-an malonil-CoA eratzeko.
Acetyl CoA Carboxylase Entzima Jarduera
ondorengo guztien abiadura zehazten du
sintesi erreakzioak lc
citrate entzima aktibatzen du zitosoletan
acetil CoA carboxylase. Malonyl CoA-n
Era berean, goiko transferentzia inhibitzen du
gantz-azidoak zitosoletik matrizera
mitokondriak inhibitzeko jarduera
kanpo azetiloa CoA: carnitina aciltransferasa,
horrela, goi-mailako oxidazioa desaktibatzen da
gantz-azidoak.
Acetil-CoA Oxaloacetate →
HS-CoA Citrate

HSCOA ATP Citrate → Acetyl-CoA ADP Pi Oxaloacetate
Azetil-CoA
zitoplasman hasierako substratu gisa balio du
lcd eta oxaloacetatoen sintesia egiteko
zitosolak eraldaketak izaten ditu
horren emaitza piruvatoa eratzen da.
Kolesterolaren biosintesia
Kolesterolaren biosintesia erretikulu endoplasmikoan gertatzen da. Molekulan dauden karbono atomo guztien iturria azetil-SCoA da, mitokondriratik dator citratoan, gantz azidoen sintesian gertatzen den bezala. Kolesterolaren biosintesia 18 ATP molekula eta NADPH 13 molekula kontsumitzen ditu.
Kolesterolaren formazioa 30 erreakzio baino gehiagotan gertatzen da, eta hainbat fasetan talka daitezke.
1. Azido mevalonikoaren sintesia.
Lehen bi sintesi erreakzioak ketogenesiaren erreakzioekin bat datoz, baina 3-hydroxy-3-metilglutaryl-ScoA sintesia egin ondoren, entzima sartzen da hydroxymethyl-glutaryl-ScoA reductase (HMG-SCOA reduktasa), azido mevalonikoa eratuz.
 |